FES-Forward Blog
Explore the Boundaries of Synthetic Biology with us. Stay up-to-date with the latest synthetic biology and DNA synthesis news, updates, and research.
Breaking Down Barriers in DNA Synthesis: Secondary Structures
SEP 26, 2024 │ 11 MIN READ
DNA synthesis solutions for secondary structures from hairpins to G-quadruplexes overcoming hurdles in gene synthesis, mRNA in-vitro transcription and DNA origami.
Secondary structures, such as hairpins and G-quadruplexes, play a vital role in the function and regulation of nucleic acids. However, these intricate formations can pose significant challenges in DNA and RNA synthesis, sequencing, and editing. Their propensity to form stable, non-linear shapes can interfere with molecular processes, complicating the accurate construction, cloning, and analysis of genetic materials. In this installment, we delve into the complexities of secondary structures, examining their impact on molecular biology workflows and exploring innovative strategies to overcome these obstacles.
What is DNA Secondary Structure?
Secondary structures can form when single stranded DNA has stretches of two or more bases that pair to fold into a non-linear conformation, with some regions becoming double stranded or higher order structures and other regions forming unpaired loops (Figure 1). The most recognized secondary structure is a DNA hairpin, where a stem-loop structure is formed between two complementary sequences within the sequence. DNA folding releases free energy, where the more energy released equates to more stable structures that require higher temperatures to dissociate (1,2). When ordering oligonucleotides for synthesis, most companies will analyze sequences for sites of secondary structures before final submission and calculate the melting temperature (Tm) and Gibbs free energy change (ΔG) for each. The lower the ΔG value, the higher the risk of secondary structure formations that will be rejected by manufacturers.
Why are Secondary Structures Challenging to Work With?
Strong DNA secondary structures cause issues during synthesis, sequencing, and replication because they render the DNA inaccessible to hybridization of complementary sequences and to enzymatic binding, ultimately affecting quality and yield of the target product. Sequences especially rich in GC content result in more stable structures, which causes large impacts on synthesis quality for both chemical and enzymatic synthesis (3). Ultimately, oligonucleotides containing structures with extreme thermal stability can make it difficult to deliver oligonucleotides that remain single stranded at common reaction temperatures (4).
In enzymatic DNA synthesis, the commonly used template-independent polymerase terminal deoxynucleotidyl transferase (TdT) binds to the last four nucleotides of the single stranded DNA without the ability to bind double stranded DNA (5,6). When secondary structures form on the initiator sequence or the 3’ end, the TdT cannot access the oligonucleotide for further elongation and eventually halts synthesis altogether, especially at typical elongation temperatures of 37C (7,8).
Much like the issues with TdT binding and extension, DNA secondary structures also present an accessibility problem for polymerases, leading to errors in PCR amplification. Polymerase stalling is often triggered by strong secondary structures, resulting in truncated products or skipped regions that lower the purity of the product (8, 9). The polymerase can “jump” over the secondary structure containing the inaccessible region thereby causing an internal deletion in the amplified product (10), or the structure can act as an arrest site that leads to truncated sequences (11).
This feature can also lead to challenges in DNA assembly. Typical assembly methods use linear, double stranded insertion fragments with ends that are homologous overlaps to the target plasmid or vector. If strong DNA secondary structures form in the overlapping regions, it prevents annealing and priming of the adjacent assembly fragments (12). Furthermore, complex sequences with secondary structures can be challenging to clone, where regions of complexity can often be truncated. Specialized cell lines and techniques, such as growing the cells at lower temperatures are common solutions employed today (13).
What are Common Examples of DNA Secondary Structure?
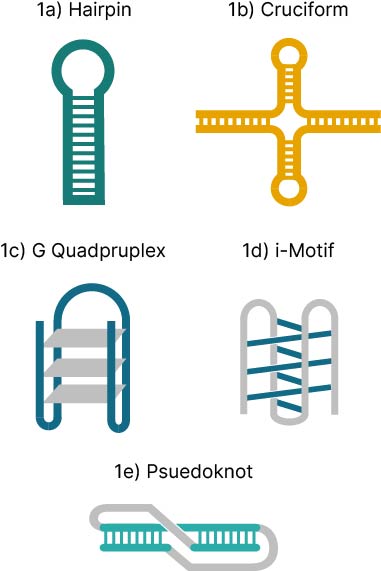
- Hairpins (aka stem-loops) (Figure 1a) are formed when self-complementary regions fold into a Watson-Crick paired stem with an intermolecular unpaired loop. These are the most common type of secondary structure, likely due to their important biological roles in regulation and their uses in molecular biology
- Protein binding sites: hairpins are enriched in regulatory regions of viral and eukaryotic genomes where they can be essential for polymerase binding and are predicted to play a role in replication, transcription, and RNA processing (14,15).
- Chromosomal functioning: long stretches of repetitive sequences can form hairpins, including homopurine tracts d(GGA)n and d(GGGA)n stabilized by unusual G-A and G-G pairs that are often found in telomeres, centromeres, and recombination hot spots (16).
- Molecular biology tools: DNA hairpins can be important components of oligonucleotide probes used to identify and capture DNA targets, in PCR suppression experiments, and DNA microarray research (17). Probes called molecular beacons function using an internally quenched fluorophore that is switched on upon binding to the target sequence have much higher specificity than linear probes and are commonly used in qPCR (18).
- Cruciform structures (Figure 1b) are induced by negative supercoiling when ≥ 6 nucleotide inverted repeats fold into a four-armed double stranded structure from two hairpin loop arms with a four-way junction (19).
- These structures are enriched near replication origins, promoters, and sites of gross chromosomal rearrangement (21) and have been shown to bind to a novel human cruciform binding protein (CBP) that may be involved in initiation of DNA replication (21).
- The human Y-chromosome is comprised mostly of inverted and direct repeats that have been highly conserved throughout evolution. It is proposed that cruciform structures within these sequences have helped stabilize and shape the chromosome functions (22).
- G–quadruplexes (G4) (Figure 1c) are formed from strong guanine self-associations (both intramolecular and intermolecular) through Hoogsteen base pair formation of guanine quartets which stack on top of each other to form structures that exhibit extreme thermal stability (4).
- Early observations of G–quadruplex structures were found in oligonucleotides encoding Tetrahymena telomeres. The stable complexes rendered the bases chemically inaccessible and were predicted to be important for telomere association and recombination (23).
- G–quadruplex structures can also be important for drug discovery, where the G4 conformation of the M08s-1 aptamer is a major contributor to its strong anticoagulant activity (24).
- G–quadruplexes are also enriched at regulatory sites within the genome, such as promoters, and serve as protein binding sites to regulate important biological processes including transcription, replication, and genome stability (25).
- Intercalated motifs (i-motifs) (Figure 1d) are four-stranded intercalated duplexes stabilized by hemi-protonated cytosine-cytosine base pairs. Like G–quadruplexes, they can form from both intermolecular associations of different DNA strands and intramolecular associations of homopolymer C runs on the same strand (26). I-motifs, also found in regulatory regions of the genome, are thought to play complementary roles with G4 structures in regulating gene expression (26). These structures are localized to the nuclei of human cells mostly during late G1 phase of the cell cycle which is the time of high transcription rates and cell growth, whereas G4s occur mostly during S phase (27).
- Pseudoknots (Figure 1e) are a more complex version of a hairpin, wherein the loop portion of a hairpin stem-loop forms Watson-Crick base pairs with additional sequence to form the pseudoknot structure (19). In terms of biological function, pseudoknots are more common in RNA and form the catalytic core of some ribozymes, telomerase, riboswitches, and work to initiate ribosomal frameshifting, a process viruses use to change reading frames to translate different mRNAs (28).
- DNA Origami is a technology for building complex nanostructures that relies on folding a long single stranded DNA scaffold into two-dimensional shapes, prepared by hundreds of designed short single stranded DNA fragments through Watson-Crick base pairing (30). The size and shape of these structures is highly customizable and has a wide range of applications including nanomaterial fabrication, nanophotonics/electronics, stabilization of enzyme cascades, molecular signaling and computation, molecular machines, drug delivery, and microscopic imaging (31).
Fully Enzymatic Synthesis (FES) Enables Successful DNA Synthesis of Secondary Structure
How can long oligos created with Fully Enzymatic Synthesis (FES) help build challenging DNA sequences containing secondary structures? Molecular Assemblies’ FES synthesis process is powerful, with high cycle efficiencies of 99.9%, enabling synthesis of up to 400mer sequences with no limitations on DNA complexity. Our proprietary TdT enzyme was intentionally designed with challenging sequences in mind by engineering the enzyme to work up to 70°C to melt secondary structures which plague most synthesis workflows, providing researchers with new, longer, more complex oligonucleotides that were previously unattainable. This provides a significant advancement to less successful previous methods to mitigate DNA secondary structure. For example, optimization of divalent metals, combined with engineering TdT for enhanced thermostability provided an improvement in oligo synthesis of hairpin structures, however the incorporation rate still lagged significantly behind non-structured synthesis (7). In another example, incorporation of modified nucleotides to destabilize the structure, fragmentation of the nucleic acid, and heat denaturation also provide benefit, albeit not to the extent of FES (29).
Figure 2 shows the synthesis quality of a sequence with 100% GC content prone to G–quadruplex formation.

To learn more about overcoming the challenges of synthesizing high DNA complexity, download our comprehensive white paper or see the first installments of this blog series on GC Content, Homopolymers, and Repetitive Sequences.
Explore More
Download the Complexity White Paper: Explore other complex DNA sequences by downloading our latest white paper on complexity.
Contact Us for More Information: Have questions or need more details? Reach out to our team for expert guidance on your DNA synthesis needs.
References
- Chen C, Wang W, Wang Z, Wei F, Zhao XS. Influence of secondary structure on kinetics and reaction mechanism of DNAhybridization. Nucleic Acids Res. 2007 May;35(9):2875–84.
- Hata H, Kitajima T, Suyama A. Influence of thermodynamically unfavorable secondary structures on DNA hybridization kinetics. Nucleic Acids Res. 2018 Jan 25;46(2):782–91.
- Molecular Assemblies Inc. Simplifying Complexity: Revolutionizing Synthesis of Challenging DNA Sequences [Internet]. 2024 [cited 2024 Sep 12]. Available from: https://molecularassemblies.com/simplifying-complexity/
- Poon K, Macgregor RB. Unusual behavior exhibited by multistranded guanine-rich DNA complexes. Biopolymers: Original Research on Biomolecules. 1998;45(6):427–34.
- Gouge J, Rosario S, Romain F, Beguin P, Delarue M. Structures of intermediates along the catalytic cycle of terminal deoxynucleotidyltransferase: Dynamical aspects of the two-metal ion mechanism. J Mol Biol [Internet]. 2013;425(22):4334–52. Available from: http://dx.doi.org/10.1016/j.jmb.2013.07.009
- Delarue M, Boulé JB, Lescar J, Expert-Bezançon N, Jourdan N, Sukumar N, et al. Crystal structures of a template-independent DNA polymerase: Murine terminal deoxynucleotidyltransferase. EMBO Journal. 2002;21(3):427–39.
- Bollum FJ. 5. Terminal Deoxynucleotidyl Transferase. In: The Enzymes. Academic Press; 1974. p. 145–71.
- Casas-Delucchi CS, Daza-Martin M, Williams SL, Coster G. The mechanism of replication stalling and recovery within repetitive DNA. Nat Commun. 2022 Dec 1;13(1).
- Voineagu I, Narayanan V, Lobachev KS, Mirkin SM. Replication stalling at unstable inverted repeats: Interplay between DNA hairpins and fork stabilizing proteins. Vol. 105, PNAS. 2008.
- Viswanathan VK, Krcmarik K, Cianciotto P. Template Secondary Structure Promotes Polymerase Jumping During PCR Amplification. Biotechniques. 1999;27:508–11.
- Jensen MA, Fukushima M, Davis RW. DMSO and betaine greatly improve amplification of GC-rich constructs in de novo synthesis. PLoS One. 2010;5(6).
- Patron NJ. DNA assembly for plant biology: Techniques and tools. Vol. 19, Current Opinion in Plant Biology. Elsevier Ltd; 2014. p. 14–9.
- Xiao S, Shiloach J, Betenbaugh MJ. Engineering cells to improve protein expression. Vol. 26, Current Opinion in Structural Biology. Elsevier Ltd; 2014. p. 32–8.
- Glucksmann MA, Markiewicz P, Malone C, Rothman-Denes LB. Specific Sequences and a Hairpin Structure in the Template Strand Are Required for N4 Virion RNA Polymerase Promoter Recognition. Vol. 70, Cell. 1992.
- Muller UR, Fitch WM. Evolutionary selection for perfect hairpin structures in viral DNA. C R Proc Soc exp Bioi Med. 1965;149:287–91.
- Huertas D, Azorín F. Structural Polymorphism of Homopurine DNA Sequences. d(GGA) n and d(GGGA) n Repeats Form Intramolecular Hairpins Stabilized by Different Base-Pairing Interactions. Biochemistry. 1996;35(40):13125–35.
- Broude NE. Stem-loop oligonucleotides: a robust tool for molecular biology and biotechnology [Internet]. Vol. 20, TRENDS in Biotechnology. 2002. Available from: http://tibtech.trends.com0167-7799/02/$-seefrontmatter
- Bonnet G, Tyagi S, Libchaber A, Kramer FR. Thermodynamic basis of the enhanced specificity of structured DNA probes. Biophysics (Oxf) [Internet]. 1999;96:6171–6. Available from: www.pnas.org.
- Baker ES, Dupuis NF, Bowers MT. DNA hairpin, pseudoknot, and cruciform stability in a solvent-free environment. Journal of Physical Chemistry B. 2009 Feb 12;113(6):1722–7.
- Bochman ML, Paeschke K, Zakian VA. DNA secondary structures: Stability and function of G-quadruplex structures. Vol. 13, Nature Reviews Genetics. 2012. p. 770–80.
- Pearson CE, Zorbas H, Price GB, Zannis-Hadjopoulos M. Inverted Repeats, Stem-Loops, and Cruciforms: Significance for Initiation of DNA Replication. Vol. 63, Journal of Cellular Biochemistry. 1996.
- Zhao J, Bacolla A, Wang G, Vasquez KM. Non-B DNA structure-induced genetic instability and evolution. Vol. 67, Cellular and Molecular Life Sciences. 2010. p. 43–62.
- Sundquist WI, Klug A. Telomeric DNA dimerizes by formation of guanine tetrads between hairpin loops. Letters to Nature. 1989;342:825–9.
- Troisi R, Napolitano V, Rossitto E, Osman W, Nagano M, Wakui K, et al. Steric hindrance and structural flexibility shape the functional properties of a guanine-rich oligonucleotide. Nucleic Acids Res. 2023 Sep 8;51(16):8880–90.
- Spiegel J, Adhikari S, Balasubramanian S. The Structure and Function of DNA G-Quadruplexes. Vol. 2, Trends in Chemistry. Cell Press; 2020. p. 123–36.
- Assi HA, Garavís M, González C, Damha MJ. I-motif DNA: Structural features and significance to cell biology. Vol. 46, Nucleic Acids Research. Oxford University Press; 2018. p. 8038–56.
- Zeraati M, Langley DB, Schofield P, Moye AL, Rouet R, Hughes WE, et al. I-motif DNA structures are formed in the nuclei of human cells. Nat Chem. 2018 Jun 1;10(6):631–7.
- Huang X, Yang Y, Wang G, Cheng Q, Du Z. Highly conserved RNA pseudoknots at the gag-pol junction of HIV-1 suggest a novel mechanism of -1 ribosomal frameshifting. RNA. 2014;20(5):587–93.
- Nguyen HK, Southern EM. Minimising the secondary structure of DNA targets by incorporation of a modified deoxynucleoside: implications for nucleic acid analysis by hybridisation. Vol. 28, Nucleic Acids Research. 2000.
Rothemund, P. W. (2006). Folding DNA to create nanoscale shapes and patterns. Nature, 440(7082), p. 297-302.
Dey, S., Fan, C., Gothelf, K.V., Li, J., Lin, C., Liu, L., Liu, N., Nijenhuis, M.A., Saccà, B., Simmel, F.C. and Yan, H., 2021. DNA origami. Nature Reviews Methods Primers, 1(1), p.13.

